
KLONOVÁNÍ GENŮ A ANALÝZA DNA
T. A. BROWN
Faculty of Life Sciences
University of Manchester, UK
Překlad Martin Fellner a kol.
Univerzita Palackého v Olomouci
ČÁST 1 ZÁKLADNÍ PRINCIPY KLONOVÁNÍ GENŮ A ANALÝZY DNA . . . 1
1 Proč jsou klonování genů a analýza DNA důležité . . . . . . . . . . . . 3
1.1 Raný rozvoj genetiky . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .3
1.2 Počátky klonování genů a polymerázové řetězové reakce (PCR) .. 4
1.3 Co je klonování genů? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
1.4 Co je PCR? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6
1.5 Proč jsou klonování genů a PCR tak důležité . . . . . . . . . . . . . . . . . . . . . . . 8
1.5.1 Izolace genu klonováním . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
1.5.2 Izolace genu pomocí PCR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .9
1.6 Jak se v knize orientovat . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . 12
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .13
2 Vektory používané při klonování genů: plazmidy a bakteriofágy . . . . . . . . . . . . . .. 14
2.1 Plazmidy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.1.1 Charakteristika plazmidů . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.1.2 Velikost a počet kopií . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
2.1.3 Konjugace a kompatibilita. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
2.1.4 Klasi. kace plazmidů . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.1.5 Plazmidy v jiných organizmech, než jsou bakterie . . . . . . . . . . . . . . . . . . 19
2.2 Bakteriofágy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
2.2.1 Základní znaky bakteriofágů . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .19
2.2.2 Lyzogenní fágy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
2.2.3 Viry jako klonovací vektory pro další organizmy . . . . . . . . . . . . . . . . . . . 27
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .27
3 Puri. kace DNA z živých buněk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .28
3.1 Izolace celkové buněčné DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .28
3.1.1 Růst bakteriální kultury a sedimentace bakteriálních buněk . . . . . . . . . . . . . . 29
3.1.2 Příprava bakteriálního extraktu . . . . . . . . . . . . . . . . . . . . . . . . .. . .30
3.1.3 Izolace DNA z buněčného extraktu . . . . . . . . . . . . . . . . . . . . . . . . . . .33
3.1.4 Zahuštění vzorku DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
3.1.5 Měření koncentrace DNA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .37
3.1.6 Izolace celkové buněčné DNA z jiných organizmů než bakterií . . . . . . . . . . . . . 38
3.2 Izolace plazmidové DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2.1 Separace na základě velikosti . . . . . . . . . . . . . . . . . . . . . . . . . . . . .41
3.2.2 Separace na základě konformace . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.2.3 Ampli. kace plazmidu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
3.3 Izolace bakteriofágové DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
3.3.1 Růst kultur k získání velkého množství částic bakteriofága . . . . . . . . . . . . . 48
3.3.2 Příprava nelyzogenního fága . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .49
3.3.3 Sedimentace fága z in. kované kultury . . . . . . . . . . . . . . . . . . . . . . . . .50
3.3.4 Izolace DNA z fágových částic . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
3.3.5 Obtíže spojené s puri. kací DNA z M13 . . . . . . . . . . . . . . . . . . . . . . . . .52
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .53
4 Manipulace s puri. kovanou DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
4.1 Enzymy používané k manipulacím s DNA . . . . . . . . . . . . . . . . . . . . . . . . . . 55
4.1.1 Nukleázy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
4.1.2 Ligázy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
4.1.3 Polymerázy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
4.1.4 Enzymy modi. kující DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .60
4.1.5 Topoizomerázy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .61
4.2 Enzymy speci. cky štěpící DNA – restrikční endonukleázy . . . . . . . . . . . . . . . . .61
4.2.1 Objev a funkce restrikčních endonukleáz . . . . . . . . . . . . . . . . . . . . . . . .63
4.2.2 Restrikční endonukleázy typu II štěpí DNA ve speci. ckých nukleotidových sekvencích.. .64
4.2.3 Tupé a kohezní konce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.2.4 Frekvence rozpoznávacích sekvencí v DNA molekule . . . . . . . . . . . . .. . . . . . 65
4.2.5 Provádění restrikční reakce v laboratoři . . . . . . . . . . . . . . . . . . . . . . . 68
4.2.6 Analýza výsledku štěpení restrikční endonukleázou . . . . . . . . . . . . . . . . . . 70
4.2.7 Zjištění velikosti molekul DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
4.2.8 Mapování pozic různých restrikčních míst na molekule DNA. . . . . . . . . . . . . . . .76
4.3 Ligace – spojování molekul DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78
4.3.1 Působení DNA ligázy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .78
4.3.2 Kohezní konce zvyšují účinnost ligace . . . . . . . . . . . . . . . . . . . . . . . . 78
4.3.3 Připojení kohezních konců na tupé konce molekuly . . . . . . . . . . . . . . . . . . . 80
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .86
5 Přenos DNA do živých buněk . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
5.1 Transformace – příjem molekuly DNA bakteriálními buňkami . . . . . . . . . . . . . . . . 90
5.1.1 Ne všechny druhy bakterií jsou stejně schopné přijímat DNA. . . . . . . . . . . . . . 90
5.1.2 Příprava kompetentních buněk E. coli . . . . . . . . . . . . . . . . . . . . . . . . . 90
5.1.3 Selekce transformovaných buněk . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
5.2 Identi. kace rekombinantů . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .93
5.2.1 Výběr buněk transformovaných plazmidem pBR322 – inserční inaktivace genu pro rezistenci k antibiotiku . .93
5.2.2 Inserční inaktivace nemusí být vždy spojena s rezistencí k antibiotiku . . . . .95
5.3 Přenos fágové DNA do bakteriální buňky . . . . . . . . . . . . . . . . . . 97
5.3.1 Transfekce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .98
5.3.2 Příprava klonovacích vektorů . in vitro . . . . . . . . . . . . . .98
5.3.3 Fágová infekce se na agarovém médiu objeví ve formě plaků . . . . . . . . . .99
5.4 Identi. kace rekombinantních fágů . . . . . . . . . . . . . . . . . . . . . . 101
5.4.1 Inserční inaktivace genu lacZ" pomocí fágového vektoru .. . . . . . . . . 101
5.4.2 Inserční inaktivace genu cI fága . . . . . . . . . . . . . . . . . . 102
5.4.3 Selekce pomocí fenotypu Spi . . . . . . . . . . . . . . . . . . . . 103
5.4.4 Selekce na základě velikosti genomu . . . . . . . . . . . . . . 103
5.5 Přenos DNA do nebakteriálních buněk . . . . . . . . . . . . . . . . . . . 103
5.5.1 Transformace jednotlivých buněk . . . . . . . . . . . . . . . . 103
5.5.2 Transformace celých organizmů . . . . . . . . . . . . . . . . . 106
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106
6 Klonovací vektory pro E. coli . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107
6.1 Klonovací vektory založené na plazmidech z E. coli . . . . . . . . 108
6.1.1 Názvosloví plazmidových klonovacích vektorů . . . . . . 108
6.1.2 Užitečné vlastnosti pBR322 . . . . . . . . . . . . . . . . . . . . . 108
6.1.3 Původ pBR322 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 109
6.1.4 Důmyslnější plazmidové klonovací vektory z E. coli . . 11
6.2 Klonovací vektory založené na bakteriofágu M13 . . . . . . . . . . 114
6.2.1 Vývoj klonovacích vektorů M13 . . . . . . . . . . . . . . . . . . 115
6.2.2 Hybridní plazmidové vektory M13 . . . . . . . . . . . . . . . .120
6.3 Klonovací vektory založené na bakteriofágu . . . . . . . . . . . . . .120
6.3.1 Segmenty genomu . mohou být odstraněny bez narušení životaschopnosti .122
6.3.2 Přírodní selekce může být použita k izolaci . . . . . . . . . . . .123
6.3.3 Inserční a substituční vektory . . . . . . . . . . . . . . . . . . . .123
6.3.4 Klonovací experimenty s inserčními nebo substitučními vektory .126
6.3.5 Velké fragmenty DNA mohou být klonovány pomocí kosmidu . 127
6.4 . Jiné velkokapacitní vektory umožňují konstrukci genomových knihoven .129
6.5 Vektory pro jiné bakterie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131
7 Klonovací vektory pro eukaryotní organizmy . . . . . . . . . . . . . . . . . . . 132
7.1 Vektory pro kvasinky a jiné houby . . . . . . . . . . . . . . . . . . . . . . . 132
7.1.1 Selekční markery pro 2µm plazmid . . . . . . . . . . . . . . . 132
7.1.2 Vektory založené na 2µm plazmidu – kvasinkové epizomální plazmidy . 133
7.1.3 YEp se může integrovat do chromozomální DNA kvasinky . . . . . . . . . 135
7.1.4 Jiné typy kvasinkových klonovacích vektorů . . . . . . . . 135
7.1.5 Umělé chromozomy můžeme použít ke klonování velkých částí DNA v kvasinkách . 138
7.1.6 Vektory pro jiné kvasinky a houby . . . . . . . . . . . . . . . . 140
7.2 Klonovací vektory pro vyšší rostliny . . . . . . . . . . . . . . . . . . . . . 141
7.2.1 Agrobacterium tumefaciens – nejmenší genový inženýr přírody . . . . . . . . . . . . . 141
7.2.2 Klonování genů v rostlinách přímým přenosem genu . . 147
7.2.3 Pokusy s použitím rostlinných virů jako klonovacích vektorů . . . . . . . . . . . . . . . . . 150
7.3 Klonovací vektory pro živočichy . . . . . . . . . . . . . . . . . . . . . . . . 151
7.3.1 Klonovací vektory pro hmyz . . . . . . . . . . . . . . . . . . . . . 151
7.3.2 Klonování u savců . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156
8 Jak lze získat klon určitého genu . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
8.1 Otázka selekce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
8.1.1 Požadovaný klon získáme pomocí dvou základních strategií .. . . . . . . . . . . . . . . . . 158
8.2 Přímá selekce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160
8.2.1 Možnosti přímé selekce rozšiřuje technika marker rescue . . . . . . . . . . . . . . . . . . 162
8.2.2 Možnosti a omezení metody marker rescue . . . . . . . . . 163
8.3 Identi. kace klonu z genové knihovny . . . . . . . . . . . . . . . . . . . . 163
8.3.1 Genomové knihovny . . . . . . . . . . . . . . . . . . . . . . . . . . . 164
8.3.2 Všechny geny nejsou exprimovány současně . . . . . . . . 165
8.3.3 mRNA lze klonovat jako komplementární DNA . . . . . . . . . . . 166
8.4 Metody identi. kace klonu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 166
8.4.1 Komplementární vlákna nukleových kyselin k sobě hybridizují . .. . . . . . . . . . . . . 166
8.4.2 Hybridizační screening kolonií a plaků . . . . . . . . . . . . 168
8.4.3 Příklady praktického využití hybridizačního screeningu 172
8.4.4 Identi. kační metody založené na detekci translačního produktu klonovaného genu . . . 177
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180
9 Polymerázová řetězová reakce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 181
9.1 Nástin polymerázové řetězové reakce . . . . . . . . . . . . . . . . . . . . 181
9.2 Detailní popis PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
9.2.1 Navržení oligonukleotidových primerů pro PCR . . . . . 184
9.2.2 Stanovení správné teploty . . . . . . . . . . . . . . . . . . . . . . . 186
9.2.3 Studium produktů PCR . . . . . . . . . . . . . . . . . . . . . . . . . 189
9.3 Problémy spojené s chybováním Taq polymerázy . . . . . . . . . . . 193
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 195
ČÁST 2 VYUŽITÍ KLONOVÁNÍ GENŮ A ANALÝZY DNA VE VÝZKUMU . . . 197
10 Studium polohy a struktury genu . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199
10.1 Jak lze studovat polohu genu . . . . . . . . . . . . . . . . . . . . . . . . . . .199
10.1.1 Lokalizace genu na malé molekule DNA . . . . . . . . . . 199
10.1.2 Nalezení polohy genu na velké molekule DNA . . . . . .202
10.2 Sekvenování DNA – zjišťování struktury genu . . . . . . . . . . . . .207
10.2.1 Sanger-Coulsonova metoda – nukleotidy ukončující řetězce . .. . . .207
10.2.2 Automatizované sekvenování DNA . . . . . . . . . . . . . . . 211
10.2.3 Sekvenování PCR produktů . . . . . . . . . . . . . . . . . . . . . 213
10.2.4 Maxam-Gilbertova metoda – chemická degradace DNA . . .. . . . . 214
10.2.5 Tvorba dlouhých sekvencí DNA . . . . . . . . . . . . . . . . . . 216
10.2.6 Úspěchy v sekvenování DNA . . . . . . . . . . . . . . . . . . . . 218
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 218
11 Zkoumání exprese a funkce genu . . . . . . . . . . . . . . . . . . . . . . . . . . . .220
11.1 Zkoumání transkriptů klonovaného genu . . . . . . . . . . . . . . . . .220
11.1.1 Elektronová mikroskopie molekul nukleových kyselin . 221
11.1.2 Analýza DNA-RNA hybridů pomocí nukleázy . . . . . .223
11.1.3 Analýza transkriptu pomocí prodloužení primeru. . . .224
11.1.4 Další techniky zkoumání RNA transkriptů . . . . . . . . .225
11.2 Zkoumání regulace genové exprese . . . . . . . . . . . . . . . . . . . . . .230
11.2.1 Identi. kace vazebných míst pro proteiny
na molekule DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 231
11.2.2 Identi. kace kontrolní sekvence pomocí
deleční analýzy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .236
11.3 Identi. kace a studium translačního produktu
klonovaného genu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .239
11.3.1 Metody HRT a HART umožňují identi. kovat
translační produkt klonovaného genu . . . . . . . . . . . . .239
11.3.2 Analýza proteinů pomocí mutageneze in vitro . . . . . . .240
11.3.3 Studium interakcí protein-protein . . . . . . . . . . . . . . . . . 247
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .250
12 Studium genomů . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .252
12.1 Genomika – jak sekvenovat genom . . . . . . . . . . . . . . . . . . . . . .253
12.1.1 Strategie „shotgun“ v sekvenování genomu . . . . . . . . .254
12.1.2 Strategie „kontigu“ klonů . . . . . . . . . . . . . . . . . . . . . . . 257
12.1.3 Použití map k sestavení sekvence . . . . . . . . . . . . . . . . . 261
12.2 Post-genomika – snaha porozumět sekvenci genomu . . . . . . . .265
12.2.1 Identi. kace genů v sekvenci genomu . . . . . . . . . . . . . .265
12.2.2 Určení funkce neznámého genu . . . . . . . . . . . . . . . . . .268
12.3 Studie transkriptomu a proteomu . . . . . . . . . . . . . . . . . . . . . . .269
12.3.1 Studium transkriptomu . . . . . . . . . . . . . . . . . . . . . . . . .270
12.3.2 Studium proteomu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 274
ČÁST 3 VYUŽITÍ KLONOVÁNÍ GENŮ A ANALÝZY DNA V BIOTECHNOLOGII . . 275
13 Produkce proteinů z klonovaných genů . . . . . . . . . . . . . . . . . . . . . . . 277
13.1 Speciální vektory pro expresi cizích genů v E. coli . . . . . . . . . .279
13.1.1 Promotor jako rozhodující složka expresního vektoru .282
13.1.2 Kazety a genové fúze . . . . . . . . . . . . . . . . . . . . . . . . . . .285
13.2. Obecné problémy spojené s produkcí rekombinantních proteinů v E. coli . ..287
13.2.1 Problémy vyplývající ze sekvence cizího genu . . . . . . .288
13.2.2 Problémy způsobené E. coli . . . . . . . . . . . . . . . . . . . . . . 291
13.3 Produkce rekombinantního proteinu eukaryotními buňkami . .292
13.3.1 Rekombinantní proteiny z kvasinek a lamentních hub . . . . . . . . . . .292
13.3.2 Využití živočišných buněk v produkci rekombinantních proteinů . . . . . . .295
13.3.3 Farming – rekombinantní protein z živočichů a rostlin . . . . . . . . . . . .297
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .300
14 Klonování genů a analýza DNA v medicíně . . . . . . . . . . . . . . . . . . . .302
14.1 Produkce rekombinantních léčiv . . . . . . . . . . . . . . . . . . . . . . . .302
14.1.1 Rekombinantní inzulin . . . . . . . . . . . . . . . . . . . . . . . . .302
14.1.2 Syntéza lidských růstových hormonů v E. coli . . . . . . . 305
14.1.3 Rekombinantní koagulační faktor VIII . . . . . . . . . . . . .306
14.1.4 Syntéza dalších rekombinantních lidských proteinů . .309
14.1.5 Rekombinantní vakcíny . . . . . . . . . . . . . . . . . . . . . . . . .309
14.2 Identi. kace genů zodpovědných za onemocnění lidského organizmu . . . . 314
14.2.1 Jak identi. kovat gen zodpovědný za genetickou chorobu . . . . . . . . . . . 315
14.3 Genová terapie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 319
14.3.1 Genová terapie dědičných chorob . . . . . . . . . . . . . . . . 319
14.3.2 Genová terapie a rakovina . . . . . . . . . . . . . . . . . . . . . . . 321
14.3.3 Etické otázky spojené s genovou terapií . . . . . . . . . . . .322
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .322
15 Klonování genů a analýza DNA v zemědělství . . . . . . . . . . . . . . . . . .323
15.1 Strategie přidání genu v rostlinném genovém inženýrství . . . . .324
15.1.1 Rostliny vytvářející vlastní insekticidy . . . . . . . . . . . . .324
15.1.2 Plodiny rezistentní k herbicidům . . . . . . . . . . . . . . . . . 331
15.1.3 Další projekty využívající strategie přidání genu . . . . .334
15.2 Odstranění genu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .334
15.2.1 Podstata antisense technologie . . . . . . . . . . . . . . . . . . .335
15.2.2 Antisense RNA a regulace dozrávání plodů rajčat . . .336
15.2.3 Další příklady použití antisense RNA v rostlinném genetickém inženýrství ..340
15.3 Problémy s geneticky modi. kovanými rostlinami . . . . . . . . . . . 341
15.3.1 Obavy spojené s bezpečností selekčních markerů . . . . 341
15.3.2 Technologie terminátoru . . . . . . . . . . . . . . . . . . . . . . . .342
15.3.3 Nebezpečí dopadu na životní prostředí . . . . . . . . . . . .344
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .345
16 Klonování genů a analýza DNA ve forenzním lékařství. . . . . . . . . . .346
16.1 Analýza DNA při identi. kaci osob podezřelých ze spáchání trestného činu . .346
16.1.1 Genetický . ngerprinting pomocí hybridizačního screeningu . .. . . . . . . . . . 347
16.1.2 Zjištění pro. lu DNA pomocí PCR krátkých tandemových repetic . . . . . . 347
16.2 Vyšetřování příbuzenských vztahů zjišťováním pro. lu DNA .350
16.2.1 Pokrevní příbuzní mají podobné pro. ly DNA . . . . . . .350
16.2.2 Zjišťování pro. lu DNA a ostatky rodu Romanovců . .350
16.3 Určení pohlaví pomocí analýzy DNA . . . . . . . . . . . . . . . . . . . .353
16.3.1 PCR sekvencí speci. ckých pro chromozom Y . . . . . .353
16.3.2 PCR genu kódujícího amelogenin . . . . . . . . . . . . . . . .354
16.4 Archeogenetika – využití DNA ke studiu evoluce člověka . . . .356
16.4.1 Kořeny moderního člověka . . . . . . . . . . . . . . . . . . . . . .356
16.4.2 DNA může být rovněž použito ke studiu prehistorických migrací lidstva . . . .359
Doplňující literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361
Slovníček . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .363
Rejstřík . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 381
Ukázka (výňatek z 1. a 8. kapitoly)
Proč jsou klonování genů a analýza DNA důležité
V polovině 19. století Gregor Mendel formuloval zákony, jimiž vysvětlil dědičnost biologických znaků. Tyto zákony vycházely z předpokladu, že dědičné vlastnosti organismu řídí faktor zvaný gen, tj. částice fyzické podstaty, která je přítomna někde v buňce. Znovuobjevení Mendlových zákonů v roce 1900 je považováno za zrod genetiky, vědy, která se snaží porozumět tomu, co jsou geny a jak fungují.
1.1 Raný rozvoj genetiky
V prvních třiceti letech existence se tato nová věda rozvíjela překvapivě rychle. S myšlenkou, že geny jsou lokalizovány na chromozomech, přišel v roce 1903 W. Sutton a experimentálně ji v roce 1910 potvrdil T. H. Morgan. Morgan a jeho spolupracovníci následně rozvíjeli postupy mapování genů (gene mapping) a do roku 1922 předložili komplexní analýzu relativních poloh více než 2000 genů čtyř chromozomů octomilky obecné, Drosophila melanogaster.
Aniž bychom chtěli snižovat brilantnost těchto klasických genetických studií, faktem zůstává, že molekulární podstata genu nebyla zřejmá až do 40. let 20. století. Teprve na základě experimentů Averyho, MacLeoda a McCartyho z roku 1944 a experimentů Hersheyeho a Chaseové z roku 1952 byla přijata koncepce, že deoxyribonukleová kyselina (DNA) představuje genetický materiál: do té doby se obecně předpokládalo, že funkci genů mají proteiny.
Poznání úlohy DNA bylo pro genetický výzkum obrovským stimulem a ke zlatému věku genetiky, v pořadí již druhému, přispěla celá řada slavných biologů (k nejvýznamnějším patřili Dellbrück, Charga. , Crick a Monod). Během 14 let, mezi léty 1952 a 1966, byla objasněna struktura DNA, rozluštěn genetický kód a byly popsány procesy transkripce a translace.
1.2 Počátky klonování genů a polymerázové řetězové reakce (PCR)
Po letech aktivity a objevování následovalo období klidu, zklamání, kdy se někteří molekulární biologové (jak se nazvala nová generace genetiků) domnívali, že prakticky všechno podstatné už bylo vysvětleno. Ve skutečnosti ale panovala nespokojenost, protože experimentální postupy konce 60. let minulého století nebyly dostatečně propracované a nedovolovaly gen detailně
prozkoumat.
V letech 1971 až 1973 se v genetickém výzkumu znovu začalo něco dít, a to díky tzv. revoluci v experimentální biologii. Byla vyvinuta zcela nová metodologie umožňující víceméně úspěšné projektování a realizaci dříve neproveditelných experimentů. Tyto metody, souhrnně nazývané technologie rekombinantní DNA nebo také genové inženýrství, jejichž jádrem bylo klonování genů, odstartovaly novou éru genetiky. Vedly k rychlým a účinným technikám sekvenování DNA, s jejichž pomocí bylo možné určit struktury jednotlivých genů. V 90. letech 20. století tyto postupy vyvrcholily impozantními projekty zaměřenými na sekvenování genomu včetně projektu sekvenování lidského genomu, jenž byl dokončen v roce 2000. Z těchto metod vycházely nové postupy výzkumu regulace jednotlivých genů, na jejichž základě mohli biologové usuzovat na souvislosti mezi odchylkami v regulaci genů a vznikem onemocnění, jakým je např. rakovina. Tyto techniky daly vznik moderní biotechnologii, která využívá geny k výrobě proteinů a jiných sloučenin potřebných v medicíně a v průmyslové výrobě.
V 80. letech, kdy vyvrcholil rozruch způsobený revolucí v oblasti klonování genů, bylo nepředstavitelné, že by byla na dosah další, právě tak nebývalá a převratná metoda. Mezi vědci zabývajícími se výzkumem DNA se traduje, že polymerázová řetězová reakce (polymerase chain reaction, PCR) je výsledkem náhlé inspirace, nápadu, který dostal Kary Mullis jednoho večera roku 1985 při cestě autem podél pobřeží Kalifornie. Přišel na výjimečně jednoduchou
metodu, která klonování genů ideálně doplňuje. PCR usnadnila řadu postupů, které sice bylo možné provádět i v době, kdy bylo známo pouze klonování, ale jen s obtížemi. Tato metoda rozšířila možnosti analýzy DNA a umožnila molekulárním biologům nalézt nové aplikace v oblastech vzdálených tradiční medicíně, zemědělství a biotechnologii. Archeogenetika, molekulární ekologie a analýza DNA v soudním lékařství jsou pouze tři z nových disciplín, které
se formovaly v přímé návaznosti na objev PCR a které dovolují molekulárním biologům studovat evoluci lidského druhu, klást si otázky, jak se změna životního prostředí odráží v biosféře, a pracovat s účinnými nástroji v boji proti kriminalitě. Třicet let po revoluci v oblasti genetického klonování si stále připadáme jako na horské dráze a je jisté, že vzrušení z objevů ještě dlouho
nepoleví.
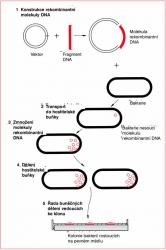
1.3 Co je klonování genů?
Základní kroky experimentu klonování jsou následující (Obr. 1.1): (1) Úsek DNA obsahující gen, který chceme klonovat, je vložen do kružnicové molekuly DNA nazývané vektor. Vznikne tak molekula rekombinantní DNA.
Kapitola 8 Jak lze získat klon určitého genu
V předchozích kapitolách jsme se zabývali základní metodologií genového klonování a podali jsme přehled vektorových typů, s nimiž se pracuje při klonování v bakteriích, kvasinkách, rostlinách a živočiších. Dále bychom se měli zaměřit na metody, kterými lze získat klon individuálního, zcela speci. ckého genu. Správný výběr představuje rozhodující zkoušku celého experimentu, protože úspěch nebo neúspěch pokusu často závisí na tom, zda lze strategii
vedoucí k přímé selekci klonů požadovaného genu, nebo alespoň jejich rozlišení od ostatních rekombinantů vůbec navrhnout. Podaří-li se tento problém vyřešit a klon získat, je možné získat informace o příslušném genu pomocí celé škály technik. Nejdůležitější z nich budou popsány v Kapitole 10 a 11.
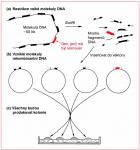
8.1 Otázka selekce
Jestliže molekulární biologové potřebují získat klon konkrétního genu, stojí před problémem znázorněným na Obr. 1.4. Dokonce i ty nejjednodušší organizmy jako např. E. coli obsahují několik tisíc genů a restrikčním štěpením celkové buněčné DNA nevznikají jen fragmenty nesoucí požadovaný gen, ale také mnoho jiných fragmentů nesoucích jiné geny (Obr. 8.1 (a)). Během ligace k selekci jednotlivých úseků nedochází: vznikají různé rekombinantní molekuly DNA obsahující různé části DNA (Obr. 8.1 (b)). Po transformaci máme širokou paletu rekombinantních klonů (Obr. 8.1 (c)), mezi kterými musíme identi. kovat ten správný.
8.1.1 Požadovaný klon získáme pomocí dvou základních strategií
Požadovaný klon lze získat celou řadou postupů, které jsou všechny variacemi
dvou postupů základních.
(1) Přímá selekce požadovaného genu (Obr. 8.2 (a)), při které klonovací pokus koncipujeme tak, abychom získali právě jen klony požadovaného genu. Selekce pravidelně probíhá po přenesení na agarové médium. (2) Identi. kace klonu pomocí genové knihovny (Obr. 8.2 (b)); začít musíme úvodním klonovacím experimentem „shotgun“ vedoucím ke vzniku knihovny klonů, která bude reprezentovat všechny geny nebo většinu genů, které se v buňce vyskytují. Analýzou jednotlivých klonů pak zjistíme ten správný.
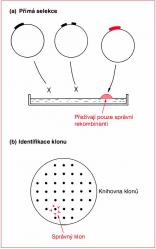
Obecně lze říci, že je upřednostňována přímá selekce, protože je rychlá a dává jednoznačné výsledky. Nemůžeme ji však použít pro všechny geny, jak si ukážeme dále. Z toho plyne, jak jsou techniky identifikace klonu důležité, a to tím spíš, že už existují kompletní genomové knihovny mnoha organizmů.
8.2 Přímá selekce
Abychom mohli provést selekci klonovaného genu, musíme transformanty nanést na agarové médium, na kterém porostou pouze požadované rekombinanty, a žádné jiné. Takto získáme kolonie, které budou obsahovat buňky s požadovanou rekombinantní molekulou DNA.
Nejjednodušší typ přímé selekce se uplatňuje tehdy, když požadovaný gen speci. kuje rezistenci k antibiotiku. Ukážeme si to na pokusu, ve kterém budeme klonovat gen pro rezistenci ke kanamycinu z plazmidu R6-5. Plazmid pR6-5 nese geny pro rezistenci ke čtyřem antibiotikům: kanamycinu, chloramfenikolu, streptomycinu a sulfonamidu. Gen pro rezistenci ke kanamycinu leží na jednom z 13 úseků EcoRI ...
Konec výňatku z knihy.
Knihu lze objednat ve Vydavatelství UP
e-mail: vup(at)upol.cz
Vlkodlaci - jak se splétají řetězce lidských a vlčích osudů
Autor: Redakce (22.02.2005)
Jak se dělá evoluce a co na to žirafí krk
Autor: Pavel Houser (14.04.2004)
Diskuze:


