Potíže se znalostmi evolučního původu křídla pramení z nedostatečného fosilního základu. Bezkřídlý hmyz je znám ze siluru (před cca 400 mil. lety), zástupci 10 různých řádů s plně vyvinutými křídly se objevují ve svrchním karbonu už před 320 mil. lety. Mezi těmito obdobími nebyla nalezena žádná fosilie, která by objasňovala, jak se vlastně hmyzí křídlo vyvinulo.
Pro pochopení evoluce hmyzího křídla je nutno vědět, z jaké struktury předka létacího hmyzu se vyvinulo, a jakou tato hypotetická struktura měla funkci. Těžko si totiž představit, že se křídlo vyvinulo jen tak z ničeho, že zničehonic vyrostlo na hrudi. Evoluce je slepá, nic neplánuje, a jen málo struktur všech známých organismů slouží svému původnímu účelu. Problém je, že žádné křídlu podobné struktury neznáme ani u korýšů, nejbližších příbuzných hmyzu, ani ostatních členovců.
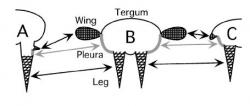
Nejen tento fakt vedl velkou část vědců 20. století k názoru, že hmyzí křídla vznikla opravdu de novo z laterálních výrůstků hřbetních destiček (terg) hrudních článků. Jako oblíbený „chybějící mezičlánek“ řetězce vedoucího k okřídlenému hmyzu byly prezentovány chvostnatky (Archeognatha), jež skutečně na hrudi mají rozšířená paranota. Nejen chvostnatky (plus někteří další, fosilní zástupci hmyzu) podporují tuto hypotézu. Pro hovoří například i spojení křídelního zárodečného pupenu (primordia) a terga u hemimetabolního hmyzu (s proměnou nedokonalou). Další „de novo“ hypotéza se například snaží vysvětlit vznik hmyzího křídla z pleury (postranní blanka, spojující hřbetní tergum a břišní sternum).
Jak jsem však již zmínil, jen málo funkčních struktur žijících organismů vzniká takříkajíc „z ničeho“. Většinou měly původně zcela jinou funkci, než v jaké je známe nyní, případně se funkčnost znásobila. Notoricky známý příklad je ptačí peří – na první pohled geniální materiál, nezbytný pro elegantní létání všech ptáků. Původní funkce byla ovšem pouze ochranná a termoizolační – stejně jako srst savců. Jiný příklad – lidská ruka, podle mnohých symbol dokonalosti. Jistě by se hodila i předku všech dnešních savců. Z původní ploutve se však nejprve vyvinula nášlapná končetina, a až u vyšších primátů se přední končetina lokomoční funkce zřekla a začala (chudák) pracovat. Sledovat evoluční původ lidské ruky je ale poměrně jednoduché – příliš se neliší od tlapky rejska či mývala, a nakonec ani ta ploutev prvních obratlovců není úplně odlišná. Hmyzí křídlo je ale oříšek.

Korýši křídla nemají, jiní členovci také ne, vznikla tedy křídla skutečně jako zcela nová struktura? A jakou měly vychlípeniny terga (či pleury) původně funkci? Mohly sloužit jako regulátory teploty – analogické struktury jsou vyvinuty u nejrůznějších taxonů živočichů (např. sloní uši či stegosauří štíty). Hypotéza vzniku hmyzích křídel de novo však na tyto a podobné otázky nedokáže uspokojivě odpovědět.
Není divu, že existuje jiný soubor hypotéz, vysvětlující vznik hmyzího křídla modifikací již existující struktury. Výhoda je zřejmá – udělat kompletně nový orgán je nepochybně složitější a náročnější, než jen pozměnit výrobní plán starého. Otázka zůstává, z čeho by se takové křídlo dalo přetvořit. Mnozí korýši mají rozvětvené končetiny. Ústní ústrojí hmyzu („hlavové končetiny“) je rozvětvené dodnes, a již zmíněné chvostnatky mají rozvětvené i končetiny kráčivé. Protože takové končetiny nalézáme i u mnoha dalších fosilních členovců, lze předpokládat, že rozvětvené končetiny měl i dávný členovec, původní předek všech korýšů a hmyzu (skupiny Pancrustacea = Tetraconata). Podle výzkumů byla noha primitivních členovců jedenáctičlenná, přičemž některé články nesly různé přívěsky – větve. Ty se nazývají exity (umístěné dorsálně) a endity (umístěné ventrálně). Další soubor hypotéz tedy předpokládá, že se křídla vyvinula z částí končetin – z jejich dorsálních přívěsků. Jedna z těchto teorií popisuje vznik hmyzího křídla z epicoxy (jeden z proximálních článků původní končetiny). Křídlo by potom bylo homologem epicoxálnímu exitu. Jiné teorie předpokládají vznik křídla z jiných částí končetiny, například z coxálního exitu (ten je zachován u chvostnatek) či exitu a enditu dohromady. Původní funkce těchto výběžků je známá – sloužily jako vnější žábry, jako dodnes u celé řady korýšů. Podle těchto teorií lze tedy s křídly homologizovat i vnější žábry larev jepic. Tyto vodní nymfy by potom vlastně měly až 7 párů křídel na zadečku. Skutečně, na první pohled nejsou žábry mnohých druhů jepic od křídel až tak odlišné.
Důkazů pro podporu hypotézy o původu křídel z částí končetin je celá řada. Nejpádněji působí výzkumy provedené na octomilkách Drosophila melanogaster. Při vývoji se část křídla i nohy odpovídajícího parasegmentu zakládají ve společném primordiu, křídelní zárodečný pupen se posléze separuje a migruje dorsálně. Pro tuto hypotézu hovoří i některé výsledky molekulární genetiky. Pokud se totiž skutečně hmyzí křídla vyvinula z exitů rozvětvené končetiny, lze předpokládat, že v těchto homologických strukturách budou zapínány i homologické geny. Při vývoji křídla u octomilek byla identifikována řada specifických genů a většina z nich je slaběji či silněji exprimována i v ostatních, kráčivých končetinách. Gen engrailed (en), ovlivňuje orientaci vzhledem k předozadní ose (anterio – posteriorní) křídla i nohy naprosto stejným způsobem. Jiné geny, např. pdm (nubbin), apterous (ap) či Distal-less (Dll) se spouští v křídlech i nohách v rozdílných oblastech. Geny pdm a ap jsou však v nohách exprimovány jen slabě, a jejich zdejší funkce je neznámá. Nabízí se tedy otázka, jaké geny jsou zapínány v exitech rozvětvené končetiny korýšů. Pro srovnání byla vybrána žábronožka Artemia franciscana a homology genů ap, pdm, en a Dll – pro přehlednost je odlišujme předponou Af. Žábronožky patří mezi lupenonožce – korýše, jejichž hrudní nožky jsou rozvětvené a mají pohybovou i dýchací funkci. V jejich žábrách by se podle této teorie měly exprimovat homologické geny podobně, jako v křídlech octomilek. Octomilky mají dva blízce příbuzné pdm geny – pdm1 a pdm2. Gen Af-pdm, izolovaný ze žábronožky, má sekvenci přibližně ekvidistantní od pdm1 a pdm2. Lze předpokládat, že Af-pdm je funkčně ekvivalentní oboum pdm genům octomilky D. melanogaster.

Engrailed se u je korýšů skutečně exprimován stejným způsobem jako u octomilky – ve všech větvích hrudní končetiny určuje předozadní orientaci. Gen Af-Dll je taktéž spouštěn ve všech částech rozvětvené nohy. Tyto dva geny jsou tedy aktivní u octomilek ve vyvíjející se noze i křídlu, u žábronožky v celé rozvětvené hrudní nožce. Naproti tomu geny Af-ap a Af-pdm jsou aktivitou omezeny jen na epipodit – dorsální vychlípeninu proximální části končetiny. V jediné z větví korýší končetiny jsou tedy zapínány geny specifické pro vývoj hmyzího křídla. Z hlediska genové exprese jsou skutečně hmyzí křídlo a epipodit korýšů identické. Další geny, aktivní jak v hmyzích křídlech tak nohách, jsou aktivní v celé rozvětvené končetině žábronožky. Výzkum provedený i na dalších korýších, rakovi Pacifastacus leniusculus a listonožce Triops longicaudatus také ukazují na podobnost genové exprese s octomilkou. Z těchto výsledků se dá proto podporovat hypotéza, že hmyzí křídlo vzniklo z rozvětvené končetiny společného předka všech korýšů a hmyzu. Jedna z větví rozvětvené končetiny dávného členovce tak dala vzniknout velmi odlišným orgánům, jako žábry některých korýšů, křídlo hmyzu či snovací bradavky a plicní vaky pavouků.
Právě u společného předka je ale největší slabina důkazů hypotézy o původu hmyzího křídla z rozvětvené končetiny. Nutný předpoklad nejsilnějších důkazů (podobná exprese genů octomilky a žábronožky a společné primordium křídla a nohy octomilky) je totiž ten, že tyto mechanismy byly stejné i u posledního společného předka hmyzu a korýšů. Pokud by tento předek například aktivoval v epipoditu jiné geny než předchůdce ap a pdm, současný stav by vznikl konvergencí. Problém je, že molekulární genetiku a vývojovou biologii na úrovni buněk fosilií více než 300 mil. let starých studovat dost dobře nejde. Zbývá tedy studovat žijící hmyz a korýše a na základě znalostech z co největšího počtu druhů posléze odhadnout stav posledního společného předka. Skoro všechny znalosti o molekulární genetice hmyzu jsou ale odvozeny od pouhého jednoho druhu, jednoho z nejúspěšnějších organismů na zemi, octomilky D. melanogaster. Ta je mimořádně vhodná pro studium genetiky – odolná, snadno se chová, má nízkou generační doba apod., její výběr má však řadu negativ. Dvoukřídlí jsou jednou z nejodvozenějších skupin hmyzu, a tvrdit, že jakýkoli znak vysoce odvozené skupiny je stejný i pro vzdáleného předka, je přinejmenším odvážné.
Právě zárodečný vývoj u octomilek je poměrně hodně odlišný od ostatního hmyzu. U hemimetabolního hmyzu je vývoj křídel postupný proces, kdy se u prvních instarů vychlipují křídelní zárodečné pupeny z ectodermu a zvětšují v každém následujícím instaru. Hmyz s proměnou dokonalou je parafyletický taxon, a protože způsob vývoje křídla je shodný pro mnohé jeho zástupce i velmi podobný některým zástupcům Holometabola (hmyz s proměnou dokonalou), je tento způsob pravděpodobně původní pro všechen okřídlený hmyz. Octomilky mají ale způsob tvorby křídla odvozený i v rámci Holometabola, a proto společné primordium křídla a nohy nemusí být přesvědčivý důkaz jejich společného původu – například někteří blanokřídlí společné primordium nemají.
Ani genová exprese není u octomilek zcela shodná jako u ostatního hmyzu. Například Dll se ve vyvíjejících se končetinách u octomilek začíná exprimovat na místě překryvu aktivity dvou genů – wingless (wg) a decapentaplegic (dpp). Exprimovaný dpp potlačuje aktivitu Dll v dorsální části primordia, a rozděluje tak zárodečný pupen na dorsální část (křídlo) a ventrální (nohu). U potemníka Tribolium castaneum a saranče Schistocerca americana je ale situace odlišná, dpp není k potlačení aktivity Dll nutný. Oproti octomilce se v končetinách rozdílně exprimují i jiné geny, například wg nebo Ultrabithorax (Ubx), u potemníka T. castaneum a ploštice Oncopeltus fasciatus.
Co to znamená? Jsou tedy obě hypotézy chybné? Pravděpodobně ne, původ hmyzího křídla z rozvětvených končetin stále vypadá velmi důvěryhodně. S jistotou to ale ještě tvrdit nemůžeme. Pro určení původních znaků hmyzu a korýšů na molekulární úrovni ještě není dostatek dat zástupců jiných taxonů, než odvozených much a brouků. Mnohé by určitě napověděly informace o vývoji a hlavně molekulární genetice jepic, pošvatek a vážek. Tyto organismy se však chovají o poznání hůře než octomilky (odhlédneme-li od náročnosti chovu, tak rozdíl čekat na novou generaci 2 týdny anebo minimálně půl roku je obrovský). Dlouhá generační doba je ale výhodou právě při hledání původních, neodvozených znaků. Doufejme, že tato data budou během několika let k dispozici a tajemství evoluce hmyzího křídla bude s jejich pomocí definitivně rozluštěno.
Zdroje:
Developmental Biology 286 (2005): 57-77.
Nature DOI: 10.1038/385627a0
Current Biology 12 (2002): 1711 – 1716.
J. Hered DOI: 10.1093/jhered/esh064
Can J Zool 61 (1983):1618–1669.
Motýli se vyvinuli až když na Zemi byly květiny. A nebo to bylo jinak?
Autor: Josef Pazdera (14.01.2018)
Diskuze:


